Anaphase: What Happens In This Stage Of Mitosis & Meiosis?
Cells are considered the fundamental units of life, given that they are the smallest biological entities that include all of the basic attributes of living things – DNA, metabolic functions, a way to maintain chemical balance and so on. Some organisms, in fact, consist of only a single cell (e.g., bacteria). The primary function of cells, viewed from the dispassionate perspective of nature, is the same as that of their parent organisms: to make copies of themselves and pass along their genetic information to subsequent generations. This evolutionary imperative means that at any given time, almost all living cells are either dividing or carrying out processes geared toward completing the next division.
In contrast to bacteria, which account for almost all of the organisms in the prokaryote group, eukaryotes (i.e., plants, animals and fungi) are, with very rare exceptions, multicellular. They have specialized organs and tissues, and accordingly, they have widely varying kinds of cells; for example, a liver cell looks markedly different from a muscle cell under a microscope. Therefore, when these somatic (i.e., body) cells of eukaryotes divide, it is for the purpose of growth, damage repair or replacing cells that are uninjured but have simply worn out over time. The type of cell division – or specifically, division of the genetic material inside the nucleus – that is associated with these non-reproductive functions is called mitosis and includes five phases: prophase, prometaphase, metaphase, anaphase and telophase. Anaphase is perhaps the most striking and elegant, as it is the short but momentous step in which duplicated chromosomes, the bearers of eukaryotic organisms' genetic material, actually separate.
DNA Basics: The Storage of Hereditary Information
DNA Basics: The Storage of Hereditary Information
Deoxyribonucleic acid (DNA) is the genetic material of all living things on Earth. "Genetic material" refers to whatever matter at the molecular level is responsible for storing and passing on information, be it to other cells in the same organism or an entirely new organism. As you may have gleaned from watching legal dramas or following real criminal trials, DNA functions like a microscopic fingerprint; every human being's is unique, apart from that of identical twins, triplets and so on.
DNA consists of long chains of units called nucleotides. These consist of three distinct chemical components: a five-carbon sugar (deoxyribose), a phosphate group, and a nitrogenous base. The "backbone" of the DNA strand is formed by alternating sugar and phosphate groups, whereas the bases in each nucleotide are linked to the sugar portion. DNA is double-stranded, with a three-dimensional helical or "corkscrew" shape; the two strands are connected to each other at each nucleotide via their bases.
The entire key to the genetic code lies in the fact that there are four different DNA bases, adenine (A), cytosine (C), guanine (G) and thymine (T). Each nucleotide, as noted, contains only one, so a long strand of DNA can be characterized in terms of its sequence of bases as this accounts for all of the variability between DNA molecules. Each triplet of consecutive bases (e.g., AAT, CGA and so forth) codes for one of 20 of the amino acids your body makes, and the 20 different amino acids are the subunits of proteins in the same way the four different nucleotides are the subunits of DNA.
A length of DNA that includes all of the bases that carry the code for a single protein product, made elsewhere in the cell by ribosomes, is called a gene.
Chromosome Structure and Function
Chromosome Structure and Function
DNA exists in prokaryotes as a single small circular molecule. Prokaryotes are simple, and accordingly, the bacterial genome (i.e., complete collection of DNA) is small enough so that it does not need to be physically folded or re-shaped in any way to get it to fit inside the cell.
In eukaryotes, the story is vastly different. The genome is sufficiently large to require a great deal of coiling, folding and cramming to allow an amount of DNA that would otherwise reach about 2 meters in length to fit inside a space 1 or 2 microns wide, a compression factor of an astonishing 1 million or so. This is done by organizing DNA in the form of chromatin, which is a protein called histone combined with DNA itself in a roughly 2-to-1 mass ratio. Although adding mass to make something smaller on the surface makes little sense, the electrochemical properties of these histones allow DNA to be super-condensed. Furthermore, they can control the extent of this compression, because, although DNA is always highly compressed, its level of condensation varies greatly with the cell cycle.
In life, chromatin is separated into discrete piece called chromosomes. Humans have 23 distinct chromosomes, 22 of which are numbered and one of which is an unnumbered sex chromosome (X or Y); other species may have more or fewer. In somatic cells, these are found in pairs, because you get one copy of each chromosome from your mother and one from your father. Correspondingly numbered chromosomes are called homologous chromosomes (e.g., the copy of chromosome 19 you get from your father is homologous to the copy of chromosome 19 you inherit from your mother). This arrangement has critical implications in cell division, discussed shortly.
The Cell Cycle
The Cell Cycle
Somatic cells have a distinct life cycle. Two identical daughter cells are produced by mitosis, which divides the cell's DNA, and the associated cleavage of the entire cell that follows, called cytokinesis. These cells then enter a G1 (first gap) phase, in which everything inside them is replicated except for the chromosomes. In the S (synthesis) phase, the chromosomes, which to this point have existed as single copies, are replicated, producing two identical copies of (in humans) all 46 chromosomes. These are called sister chromatids and are joined at a point called the centromere, the position of which differs from chromosome to chromosome. The cell then proceeds to the G2 (second gap) phase, in which the cell verifies the accuracy of its own DNA replication (errors in chromosome reproduction, while marvelously rare, do occur). Finally, the cell enters the M (mitosis) phase, which itself is subdivided into five phases of its own.
Cell Division: Mitosis and Meiosis
Cell Division: Mitosis and Meiosis
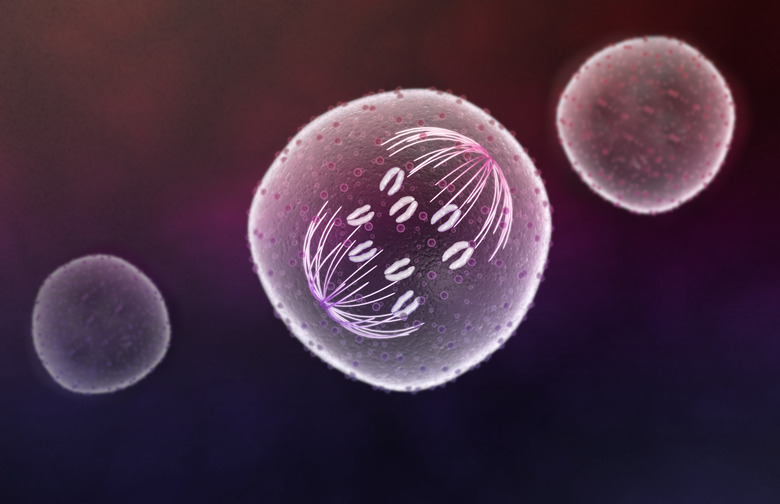
Mitosis includes five phases: prophase, prometaphase, metaphase, anaphase and telophase. Some sources combine prometaphase and metaphase into a single phase. Prophase is the longest of these and is mostly preparatory, with the nuclear membrane around the chromosomes dissolving. The chromosomes appear as highly condensed in prophase, and the spindle fibers, made of microtubules and tasked with ultimately separating the replicated chromosomes, appear. Also, two twin structures called centrosomes appear on either side of the cell, along an axis perpendicular to that along which the cell is preparing to divide.
In prometaphase, the chromosomes migrate toward the center of the cell, away from the centrosomes, while the spindle fibers extend inward as well and join the centromeres of each chromosome at a point called the kinetochore. In metaphase proper, the chromosomes line up "perfectly" along the axis of division, called the metaphase plate, with this axis passing through their centromeres. After anaphase, in which the sister chromatids are separated, comes telophase; this is a de facto reversal of prophase, with new nuclear membranes forming around the two daughter nuclei. The cell as a whole then undergoes cytokinesis.
What Happens in Anaphase?
What Happens in Anaphase?
In mitosis, anaphase is marked by the drawing apart of sister chromatids by the spindle fibers on each side of the cell. The result is the creation of daughter chromosomes. Genetically, these are identical to the sister chromatids, but the label helps emphasize the fact that new cells are soon to be formed.
In meiosis, which is the formation of gametes, or germ cells, the situation is different. Meiosis is divided into meiosis I and II, and accordingly, each of these includes its own anaphase, named anaphase I and anaphase II. In meiosis I, homologous chromosomes join to one another and form a line of 23 structures along the metaphase plate, instead of 46 individual chromosomes doing this a la mitosis. Thus in anaphase I, it is homologous chromosomes that are drawn apart, not sister chromatids, so the centromeres of the individual chromosomes remain intact. This results in daughter cells that contain 23 individual, replicated chromosomes, but these are not identical to each other thanks to exchange of material between homologous chromosomes prior to anaphase I. Each of these non-identical meiotic daughter cells then undergoes meiosis II, which is very similar to ordinary mitosis except that only 23 chromosomes are separated at their centromeres rather than 46. Thus anaphase II is functionally almost indistinguishable from anaphase in mitosis. After telophase II, the result is a total of four gametes, each with 23 chromosomes; these are spermatocytes in human males and oocytes in females, but all eukaryotes, including plants, undergo meiosis as organisms that utilize sexual reproduction.
Anaphase A
Anaphase A
Molecular biologists have found it convenient to refer to anaphase A and anaphase B to describe the events of this phase of division. Anaphase A is the migration of chromosomes toward the centrosomes via the mechanical shortening of the microtubules serving as the connecting fibers. This is what most people with a passing familiarity with mitosis and its phases think of when "anaphase" comes to mind, for the separation of sister chromatids to generate daughter chromosomes is rapid and dramatic.
The word "kinetochore" means "movement place," and in many cells, despite the extremely tiny size of the structures within chromosomes, as well as the chromosomes themselves, the spindle fibers pulling the chromatids apart at the kinetochore can be well visualized using bright-field microscopy.
The key aspect of anaphase A is that the movement of chromatids toward the poles of the cell actually occurs in concert with the microtubules of the spindle fibers being disassembled. This appears to mean that after the spindle apparatus has provided an initial "pull" toward the poles, enough momentum is generated so that the chromatids can continue to drift poleward even as the spindle fibers begin to undergo dismantling.
Anaphase B
Anaphase B
Anaphase B may be regarded as a sort of hidden element of the anaphase process. It sometimes occurs in concert with anaphase A, whereas in other cells these two processes unfold sequentially.
In anaphase, when the chromatids are pulled apart and migrate toward the poles (sides) of the cell, the entire cell, of necessity, widens and becomes more oblong. If this did not happen, subsequent division of the nucleus would not be as neat and would result in improperly sized daughter cells. This is powered by elongation of some of the spindle fibers that extend from opposite poles and overlap in the middle, without being connected to any chromosomes. These fibers undergo cross-linking, and as a result, these cross-links "push" in a direction that moves the fibers between them in opposite directions. When you think about it, a mechanism that pulls fibers from the sides of the cells and a mechanism that pushes them apart from the middle are actually working in tandem.
Cite This Article
MLA
Beck, Kevin. "Anaphase: What Happens In This Stage Of Mitosis & Meiosis?" sciencing.com, https://www.sciencing.com/what-happens-in-anaphase-13714434/. 21 September 2018.
APA
Beck, Kevin. (2018, September 21). Anaphase: What Happens In This Stage Of Mitosis & Meiosis?. sciencing.com. Retrieved from https://www.sciencing.com/what-happens-in-anaphase-13714434/
Chicago
Beck, Kevin. Anaphase: What Happens In This Stage Of Mitosis & Meiosis? last modified August 30, 2022. https://www.sciencing.com/what-happens-in-anaphase-13714434/