Nucleus: Definition, Structure & Function (With Diagram)

The cell is the fundamental organizational and functional component in living things, being the simplest natural construct that includes all of the properties assigned to life. Indeed, some organisms consist of only a single cell.
The most outstanding visual and functional feature of a typical cell is its nucleus.
The best cell nucleus analogy is that, at least in _eukaryotes_, it is the "brain" of the cell. much the same way that a literal brain is the control center of the parent animal.
In _prokaryotes_, which have no nuclei, the genetic material sits in a characteristic loose cluster in the cytoplasm of the cell. While some eukaryotic cells are anucleate (e.g., red blood cells), most human cells contain one or more nuclei that store information, dispatch commands and perform other "higher" cell functions.
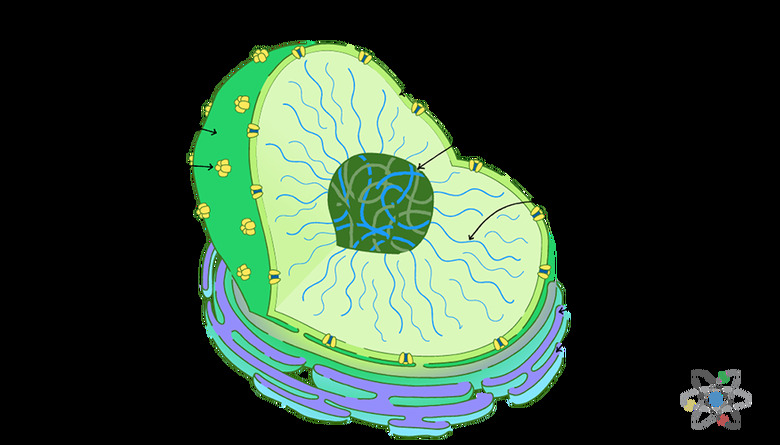
The Structure of the Nucleus
The Structure of the Nucleus
**Guarding the Fortress:** The nucleus is one of many **organelles** (French for "little organ") found in eukaryotic cells.
All cells are membrane-bound by a double membrane, usually just called the _cell membrane_; all organelles also have a double plasma membrane that separates the organelle from the **cytoplasm**, the gelatinous substance that constitutes most of the mass of a cell's interior.
The nucleus is normally the most prominent organelle when a cell is viewed under a microscope, and it is unquestionably pre-eminent in terms of importance of function.
Just as an animal's brain, though carefully shielded in as safe a physical space as possible, has to communicate with the rest of the body in various ways, the well-guarded nucleus exchanges material with the rest of the cell through a variety of mechanisms.
While the human brain is fortunate to be protected by a bony skull, the nucleus relies on a nuclear envelope for protection.
Since the nucleus is within a structure that itself is protected from the external world by a cell membrane (and in the case of plants and some fungi, a cell wall), specific threats to the nucleus should be minimal.
**Meet the Nuclear Security Team:** The nuclear envelope has the characteristics of a double plasma membrane, like that surrounding all organelles.
It contains openings called nuclear pores, through which substances can be exchanged with the cell cytoplasm in accordance with real-time requirements.
These pores actively control the transport of larger molecules, such as proteins, into and out of the nucleus proper. Smaller molecules, however, such as water, ions (e.g., calcium) and nucleic acids such as **ribonucleic acid (RNA)** and **adenosine triphosphate (ATP**, a source of energy), can pass freely back and forth through the pores.
In this way, the nuclear envelope itself, apart from its contents, contributes to the regulation of information transmitted from the nucleus to the rest of the cell.
**The Business of Nuclear Government:** The nucleus contains **deoxyribonucleic acid (DNA)** packed into coiled molecular strings called **chromatin**.
This functions as the genetic material of the cell, and the chromatin is divided in humans into 46 paired units called **chromosomes**.
Every chromosome is really nothing more than an extremely long strand of DNA along with an ample smattering of proteins called _histones_.
Finally, the nucleus also contains one or more nucleoli (singular nucleolus).
This is a condensation of DNA that codes for the organelles known as **ribosomes**. Ribosomes, in turn, are responsible for the manufacture of almost all proteins in the body. Under a microscope, the nucleolus appears dark in relation to its surroundings.
Genetic Information the Nucleus
Genetic Information the Nucleus
As noted, the fundamental molecule of the chromatin and chromosomes in the nucleus, and hence the basic molecule of genetic information, is DNA.
DNA consists of monomers called **nucleotides**, each of which in turn has three subunits: a five-carbon sugar called deoxyribose, a phosphate group and a nitrogenous base. The sugar and phosphate sections of the molecule are invariant, but the nitrogenous base comes in four types: **adenine (A), cytosine (C), guanine (G) and thymine (T).**
A single nucleotide thus contains a phosphate bonded to deoxyribose, which is bonded on its opposite side to whichever nitrogenous base is present. Nucleotides are, logically, named for the nitrogenous base they contain (e.g., A, C, G or T).
Finally, the phosphate of one nucleotide is bonded to the deoxyribose of the next, thus creating a long chain or strand of DNA.
**Getting DNA in Shape:** In nature, however, DNA is not single-stranded but double-stranded. This occurs via bonding between nitrogenous bases of adjacent strands. Critically, the types of bonds that can be formed in this arrangement are limited to A-T and C-G.
This has a variety of functional implications, one of which is that if the sequence of the nucleotides in one DNA strand are known, the sequence of the strand to which it can bind may be deduced. Based on this relationship, in double-stranded DNA, one strand is _complementary_ to the other.
Double-stranded DNA is, when undisturbed by outside factors, in the form of a **double helix**.
This means that complementary bound strands are joined by bonds between their nitrogenous bases, forming something like a ladder, and that the ends of this ladder-like construct are twisted in opposite directions from each other.
If you have seen a spiral staircase, you have in a sense seen what a DNA double helix resembles. In the nucleus, however, the DNA is very tightly packed; in fact, to function in an animal cell, every cell must contain enough DNA to reach an astonishing 6 feet if it were stretched end to end.
This is accomplished through the formation of chromatin.
Chromatin, the Cellular Efficiency Expert: Chromatin consists of DNA and proteins called histones.
Portions containing DNA alone alternate with sections containing DNA wrapped around the histones. The histone components actually consist of octets, or groups of eight. These eight subunits come in four pairs. Where the DNA meets these histone octets, it wraps itself around the histones like thread being wound around a spool.
The resulting DNA-histone complex is called a nucleosome.
The nucleosomes are wound into structures called solenoids, which are further coiled into other structures and so on; this exquisite layering of coiling and packing is what ultimately allows so much genetic information to be condensed into such a small space.
The chromatin of humans is divided into 46 distinct pieces, which are the chromosomes.
Everyone gets 23 chromosomes from each parent. 44 of these 46 chromosomes are numbered and paired, so that everyone gets two copies of chromosome 1, two of chromosome 2 and so on up to 22. The remaining chromosomes are the sex chromosomes.
A male has one X and one Y chromosome, while a female has two X chromosomes.
23 is considered the haploid number in humans, while 46 is termed the diploid number. With the exception of cells called gametes, all of a person's cells contain a diploid number of chromosomes, a single complete copy of the chromosomes inherited from each parent.
Chromatin actually comes in two types, heterochromatin and euchromatin. Heterochromatin is very tightly packed even by the standards of chromatin in general, and its DNA is not usually transcribed into RNA that codes for a functional protein product.
Euchromatin is less tightly bunched, and it is typically transcribed.
The looser arrangement of euchromatin makes it easier for the molecules that participate in transcription to access the DNA up close.
 Sciencing
Sciencing
Gene Expression and the Nucleus
Gene Expression and the Nucleus
**Transcription,** the process by which DNA is used to create a messenger RNA (mRNA) molecule, takes place in the nucleus.
This is the first step in the so-called "central dogma" of molecular biology: DNA is transcribed to make messenger mRNA, which is then _translated_ into proteins. DNA contains the genes, which are simply unique lengths of DNA that code for given proteins.
The ultimate synthesis of the protein product is what scientists mean when they mention gene expression.
At the start of transcription, the DNA double helix in the region to be transcribed becomes partially unwound, resulting in a transcription bubble. At this point, enzymes and other proteins that contribute to transcription have migrated to the region. Some of these bind to a DNA sequence of nucleotides called a promoter.
The response at the promoter site determines whether the gene "downstream" will be transcribed or whether it will be ignored.
Messenger RNA is assembled from nucleotides, which are the same as those found in DNA except for two characteristics: The sugar is ribose instead of deoxyribose and the nitrogenous base **uracil (U)** takes the place of thymine.
These nucleotides are joined to create a molecule that is almost identical to the _complementary strand_ of the DNA used as a template for transcription.
Thus a strand of DNA with the base sequence ATCGGCT would have a complementary DNA strand of TAGCCGA and an mRNA transcription product of UAGCCGU.
- Each three-nucleotide combination (AAA, AAC, etc.) carries the code for a distinct amino acid. The 20 amino acids found in the human body are what make up proteins.
- Since there are 64 possible combinations of three bases from among a total of four (4 raised to the power of 3), some amino acids have multiple codons, as they are called, associated with them. But each codon invariably codes for the same amino acid.
- Transcription errors do occur in nature, leading to mutated or incomplete protein products down the line, but overall such errors are statistically rare, and their overall impact is thankfully limited.
Once the mRNA has been fully transcribed, it moves away from the DNA upon which it was assembled.
It then undergoes splicing, which removes non-protein-coding portions of mRNA (introns) while leaving intact the protein-coding segments (exons). This processed mRNA then leaves the nucleus for the cytoplasm.
Eventually, it will encounter a ribosome, and the code it carries in the form of its base sequence will be translated into a particular protein.
Cell Division and the Nucleus
Cell Division and the Nucleus
**Mitosis** is the five-phase process (some older sources list four phases) by which a cell replicates its DNA, which means replicating its chromosomes and the structures associated with them, including the nucleus.
At the start of mitosis, the chromosomes, which to this point in the cell's life cycle have sat rather loosely in the nucleus, become far more condensed, while the nucleolus does the opposite and becomes harder to visualize; during the second of the five basic stages of mitosis, called prometaphase, the nuclear envelope disappears.
- In some species, notably the fungus yeast, the nuclear envelope remains intact throughout mitosis; this process is known as closed mitosis.
The dissolution of the nuclear envelope is controlled by the addition and removal of phosphate groups to proteins within the nucleus.
These _phosphorylation_ and dephosphorylation reactions are regulated by enzymes called kinases.
The nuclear membrane forming the envelope is reduced to an assortment of small membranous vesicles, and the nuclear pores that had been present in the nuclear envelope are taken apart.
Recall that these are not mere holes in the envelope, but channels that are actively regulated to keep certain substances from simply entering and leaving the nucleus in an uncontrolled way.
- The envelope is largely made up of proteins called lamins, and when the envelope dissolves, the lamins are depolymerized and instead exist briefly as dimers, or groups of two subunits.
During telophase, the final step in mitosis, two new nuclear envelopes form around the two sets of daughter chromosomes, and the entire cell then splits in the process of **cytokinesis** to complete the cell's division.
References
- National Center for Biotechnology Information Bookshelf: The Cell: A Molecular Approach (2nd Edition): The Nucleus During Mitosis
- Scitable by Nature Education: Chromatin
- Georgia State University: HyperPhysics: Cell Nucleus and Nuclear Envelope
- Scitable by Nature Education: The Formation of Heterochromatin and RNA Interference
- OpenTextBC: The Structure of DNA
Cite This Article
MLA
Beck, Kevin. "Nucleus: Definition, Structure & Function (With Diagram)" sciencing.com, https://www.sciencing.com/nucleus-definition-structure-function-with-diagram-13717297/. 13 March 2019.
APA
Beck, Kevin. (2019, March 13). Nucleus: Definition, Structure & Function (With Diagram). sciencing.com. Retrieved from https://www.sciencing.com/nucleus-definition-structure-function-with-diagram-13717297/
Chicago
Beck, Kevin. Nucleus: Definition, Structure & Function (With Diagram) last modified August 30, 2022. https://www.sciencing.com/nucleus-definition-structure-function-with-diagram-13717297/